水杨酸SA是一种重要的植物防御激素,在植物抵御活体营养型和半活体营养型病原体的侵染过程中发挥重要作用。在拟南芥等植物中,异分支酸合酶ICS1是病原体诱导SA生物合成的关键酶,负责反应的第一步,将分支酸异构化为SA合成前体异分支酸。ICS1特异性识别并催化异分支酸合成的结构机理仍不清楚。
近日,我院明振华课题组在Plant Physiology上发表题为“Structural basis of chorismate isomerization by Arabidopsis ISOCHORISMATE SYNTHASE1”的研究论文,揭示了拟南芥异分支酸合酶ICS1催化分支酸异构化的结构及催化机制。

该研究通过LC-HRAM-MS实验发现拟南芥、水稻、大豆和番茄等植物的异分支酸合酶ICS1可以催化底物分支酸异构化生成产物异分支酸。酶活检测表明植物ICS1的催化过程具有镁离子依赖性,拟南芥ICS1的催化活性最高,水稻ICS1活性极低,烟草ICS1蛋白没有检测到催化活性。
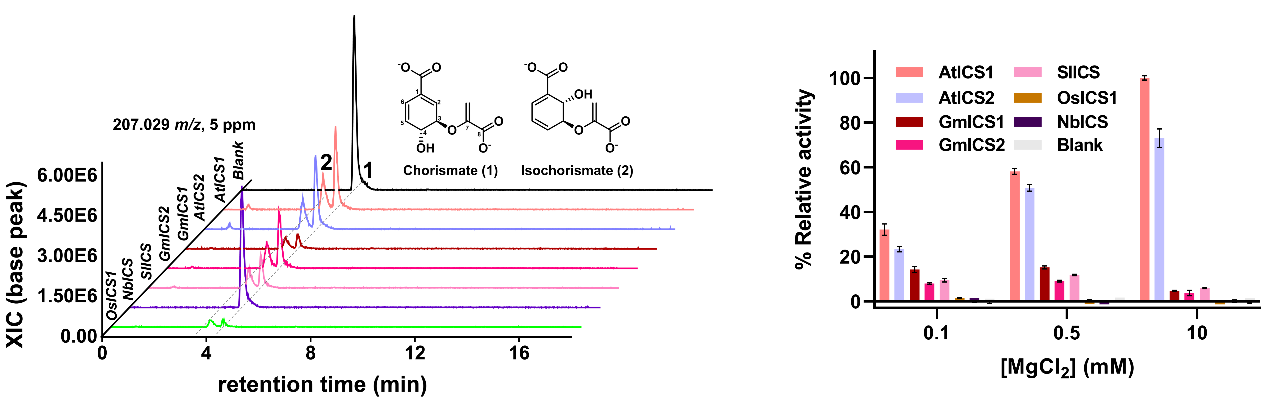
图1 植物异分支酸合酶ICS1的催化活性
为了阐明植物ICS1蛋白的催化机理,研究者解析了拟南芥ICS1及其与底物分支酸的复合物晶体结构。结构分析鉴定了ICS1用于底物识别和催化的关键氨基酸残基,突变体酶活实验表明这些氨基酸在ICS1催化过程中发挥重要作用。
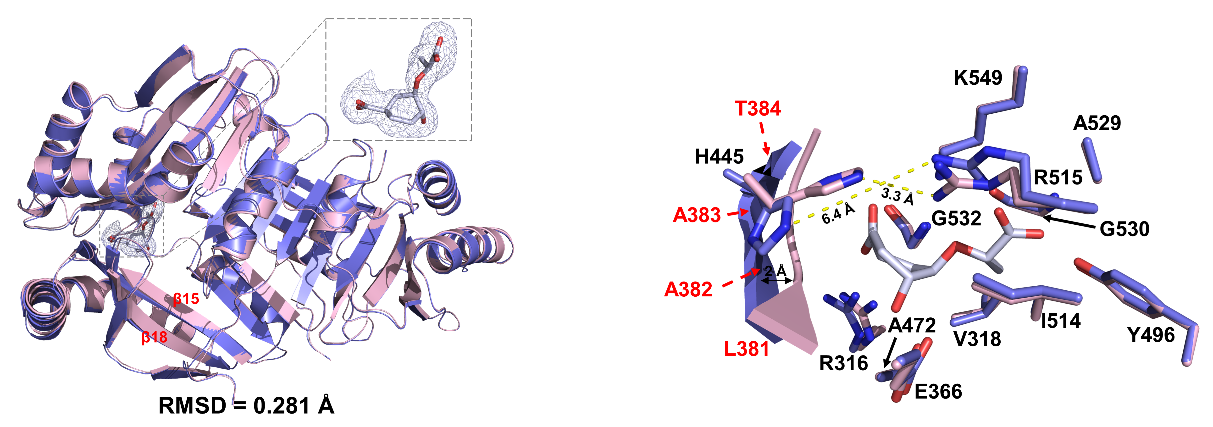
图2 ICS1识别底物的关键氨基酸残基
研究者同时发现,酶分子的催化中心与溶剂之间有一个长通道,底物穿过通道到达活性位点需要克服较高的能量屏障。分子对接和突变体实验结果表明长通道中存在一个可以结合底物的关键位点,底物与该位点的瞬时结合可以降低其穿过长通道的能量消耗,最终提升进入催化活性位点的效率。基于这些发现,研究者对低催化活性的水稻ICS1进行了理性设计,成功提高了其催化活性。
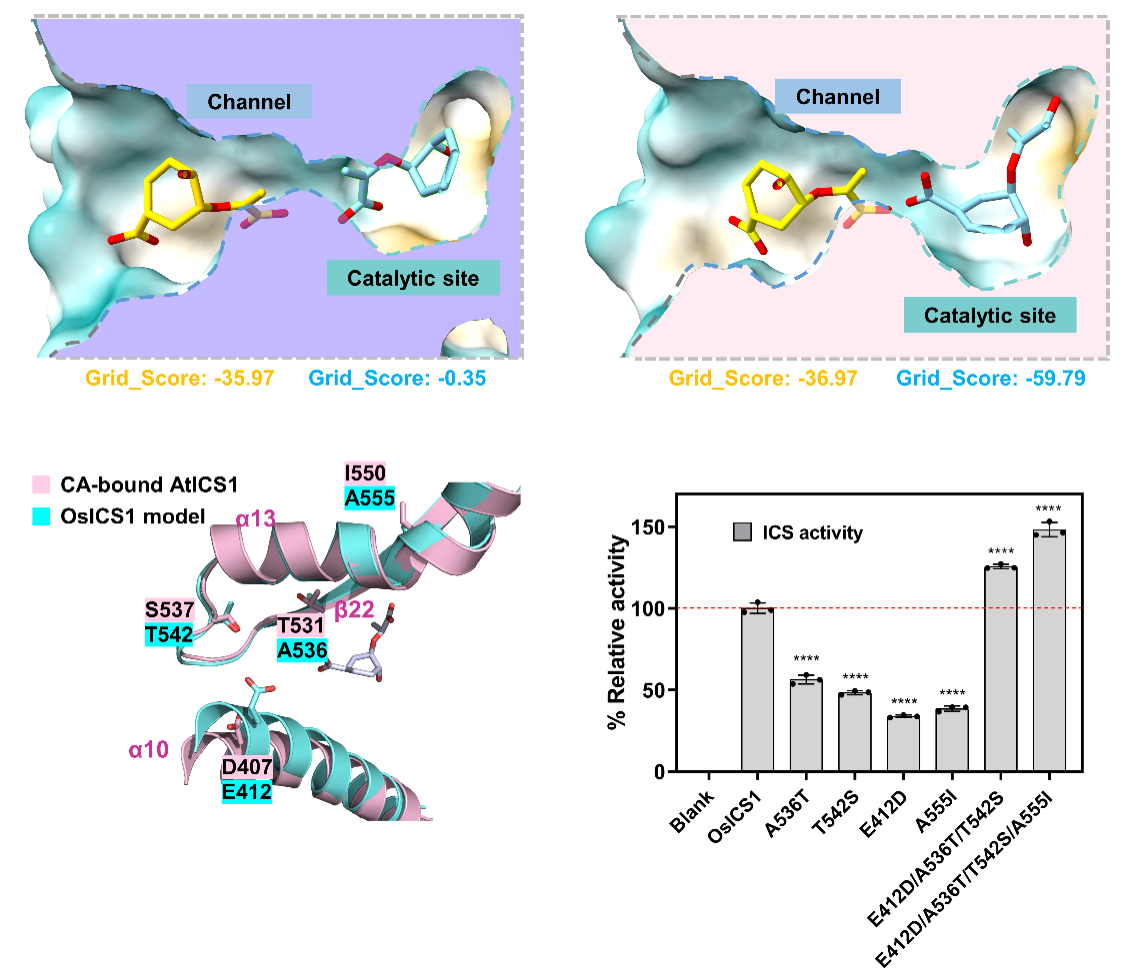
图3 ICS1通道结构瞬时结合底物以提升其递送效率
进一步的结构比对发现,植物ICS1在催化反应前可能形成一个封闭的反应口袋。研究者于是分析了这类口袋的形成条件,并提出了ICS1催化的门控调节模型:底物分支酸结合前,ICS1处于静息状态。此时参与门控调节的两个关键的谷氨酸残基相距较远,催化口袋处于开放状态。ICS1利用通道瞬时结合底物分支酸,可以帮助底物高效地穿过通道,抵达活性中心。随后镁离子协同参与门控调节的两个谷氨酸封闭通道,形成闭合的催化口袋,确保酶分子可以有效催化分支酸的异构化。产物异分支酸形成后通道重新打开,产物释放,ICS1回到最初的静止状态,完成一个催化循环。
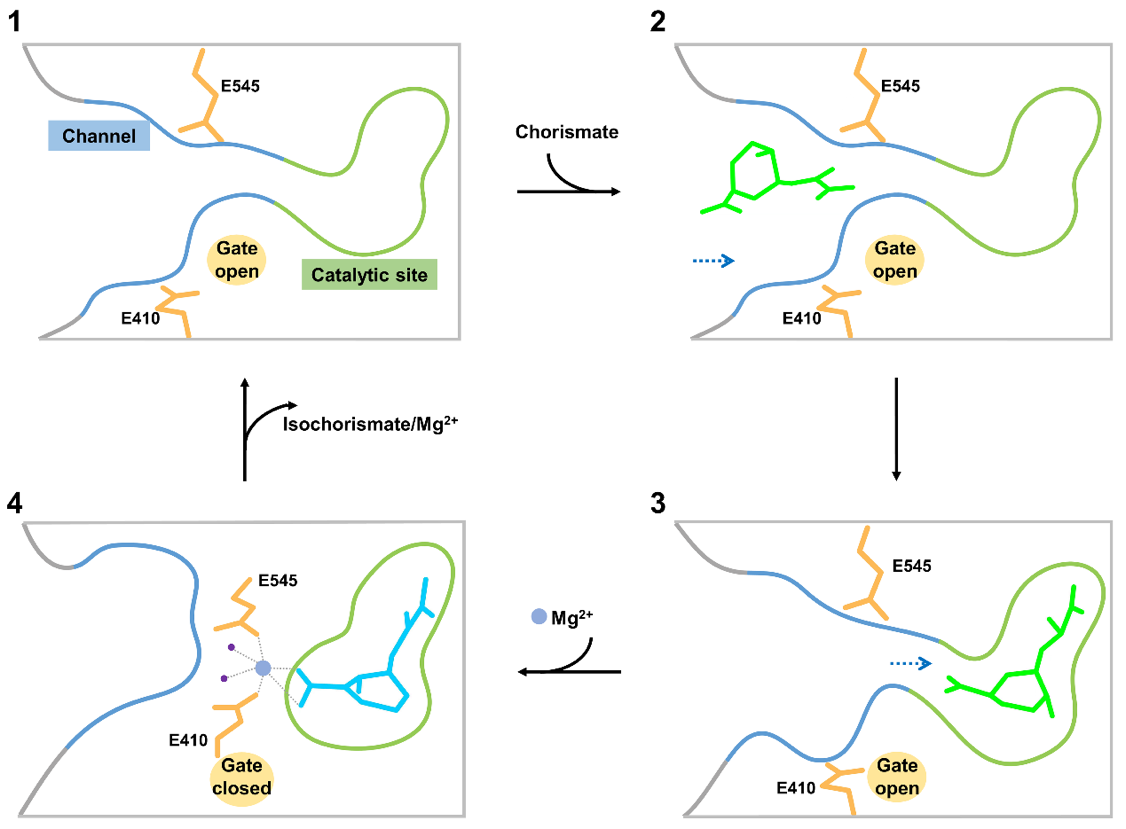
图4 ICS1催化的门控调节模型
广西大学开云在线注册,开云(中国)博士研究生苏子惠是论文第一作者,明振华教授为通讯作者。加拿大英属哥伦比亚大学张跃林教授为研究工作的顺利开展做出了重要贡献,上海同步辐射光源为衍射数据收集提供了大力支持。研究工作得到了国家自然科学基金、广西杰青基金、广西“八桂学者计划”的资助。
论文链接:
https://doi.org/10.1093/plphys/kiae260
